Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2018-07
-
CU CPT 22 All DGKs have at least two
2019-09-30
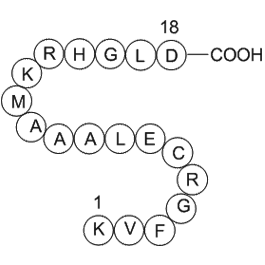
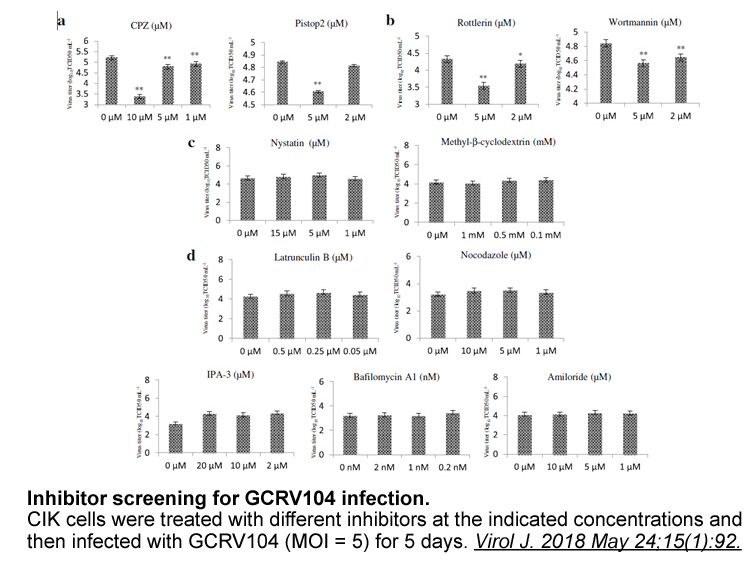
All DGKs have at least two cysteine-rich regions homologous to the C1A and C1B motifs of PKCs [26]. In theory, these domains may bind DAG, perhaps localizing DGKs to where DAG accumulates. However, no DGK C1 domain has so far been conclusively shown to bind DAG. In fact, structural predictions sugge
-
However there are still some issues that
2019-09-30
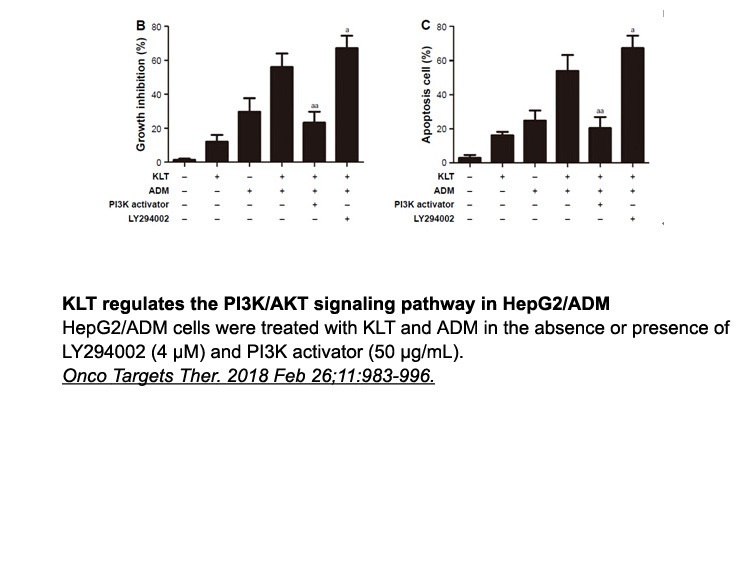
However, there are still some issues that remain poorly understood. For instance, how TGF-β modulates DDR2 expression and whether Smads participate in the regulation. How does DDR2 regulate the expression of PTHrP via Runx2? Is there any other kinase downstream of DDR2 participating in PTHrP regulat
-
The collagen binding membrane proteins discoidin domain rece
2019-09-30
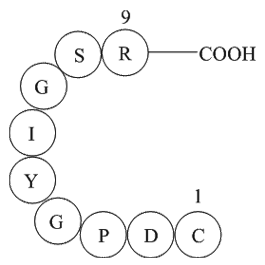
The collagen-binding membrane proteins, discoidin domain receptors 1 and 2 (DDR1 and DDR2) belong to the family of receptor tyrosine kinase and are expressed in a variety of mammalian cells.7, 8 These transmembrane glycoproteins (∼125 kDa) have been found to be over-expressed or atypically expressed
-
Our recent discovery M P H R
2019-09-30
Our recent discovery (M.P., H.R., A.S.) of a highly selective and in vivo active DDR1 small-molecule inhibitor provides evidence that DDR1 is a druggable pharmaceutical target, and some details of our efforts are provided below. To avoid the repurposing of known kinase inhibitor structural motifs,
-
The synthetic routes to the substrates based on
2019-09-29
The synthetic routes to the substrates based on the 2,3-dihydroxynaphthalene and 6,7-dibromo-2,3-dihydroxynaphthalene cores are depicted in Scheme 3. A Michael-type glycosylation of 2,3-dihydroxynaphthalene 16a gave the acetylated sugar 17a which was deprotected giving the required β-glucosidase sub
-
A prominent example of such
2019-09-29
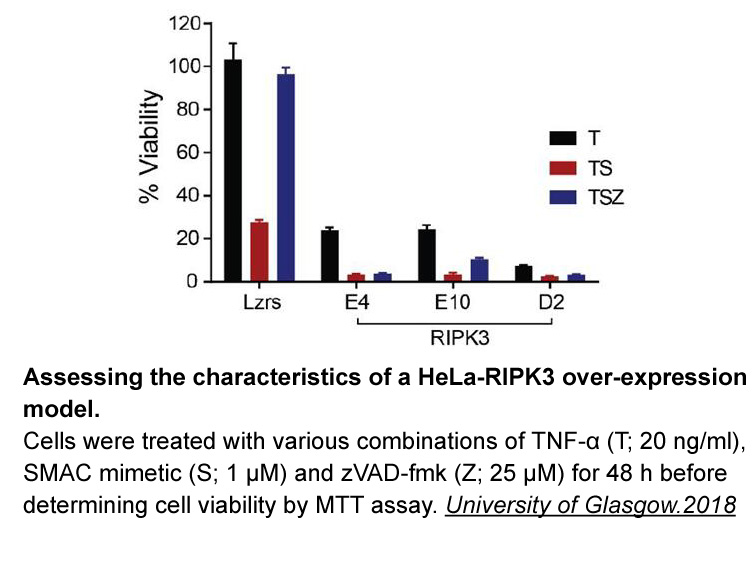
A prominent example of such enzyme is the proteasome, a highly promising target not only in cancer, but also in inflammatory and autoimmune diseases. Circulating proteasomes and respective anti-proteasome autoantibodies were detected in serum samples from patients with autoimmune diseases such as mu
-
Several eukaryotic E enzymes including BRE RNF and RAD have
2019-09-29
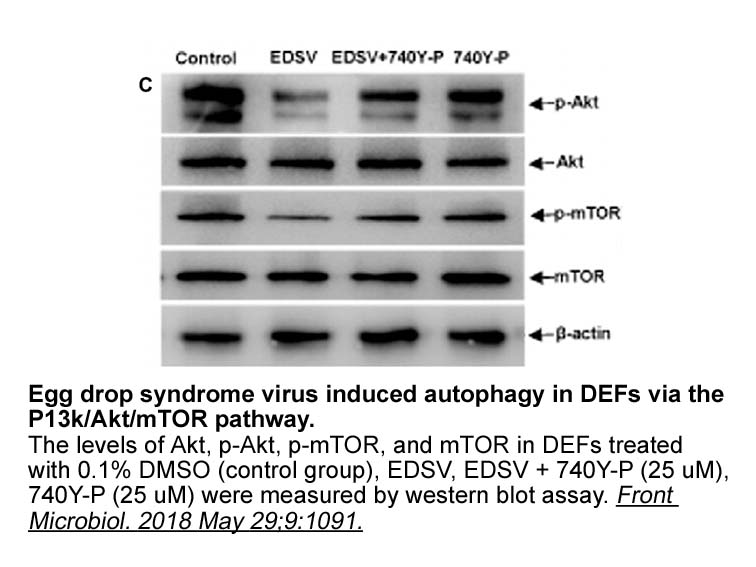
Several eukaryotic E3 enzymes, including BRE1 [49], RNF8 [50], and RAD18 [24], [51], have been reported to partner with UBE2B in the ubiquitination of various targets. We examined the expression levels of these E3 enzymes in HONE1 and TW01 cells; RAD18 was highly expressed in these 3-Deazaadenosine
-
Alpha SYN has no direct effect on forskolin induced CREB
2019-09-29
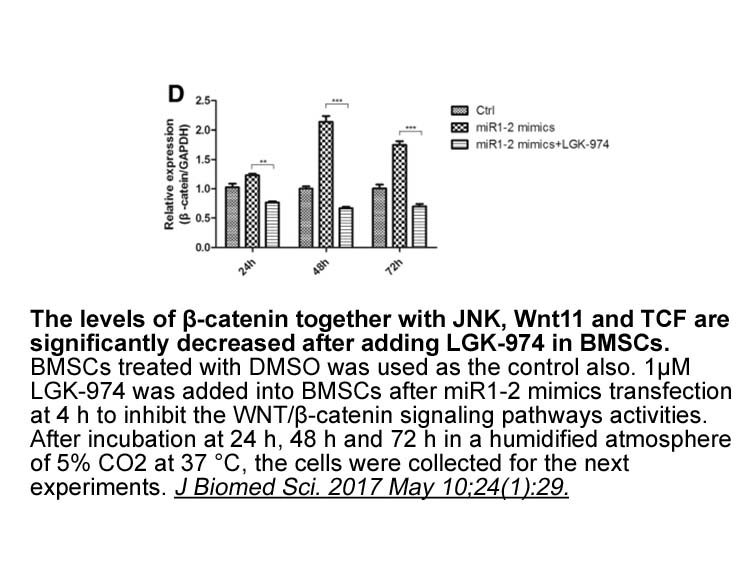
Alpha-SYN has no direct effect on forskolin-induced CREB phosphorylation, but rather blocks the stimulation of CRE-mediated transcription in the nucleus. Previously we suggested that α-SYN can enter the nucleus and is present in both cytosolic and nuclear fractions without direct interaction between
-
belinostat br Methods for assessing TLS While
2019-09-29
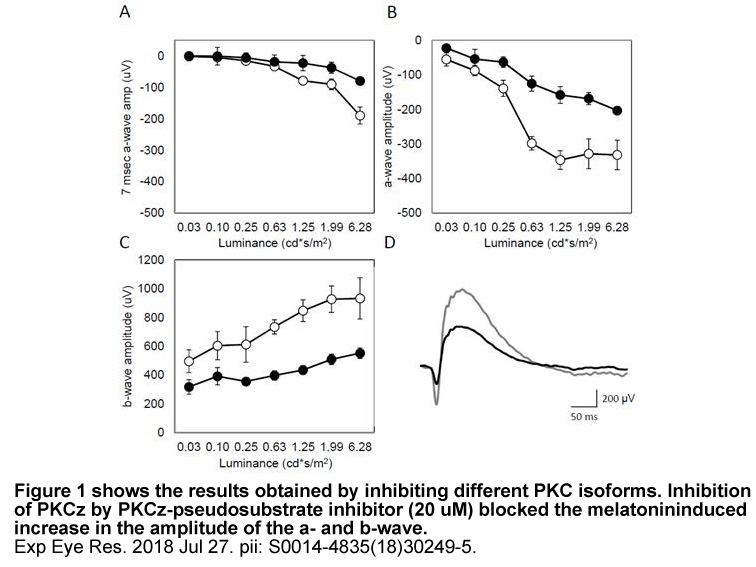
Methods for assessing TLS While the precise quantification of restricted DNA synthesis events is possible (e.g. unscheduled DNA synthesis (UDS) reveals NER), so far, it is impossible to identify TLS stretches of only a few nucleotides within the background of bulk DNA replication of normal DNA. N
-
EX 527 br Conclusions In this paper we describe
2019-09-29
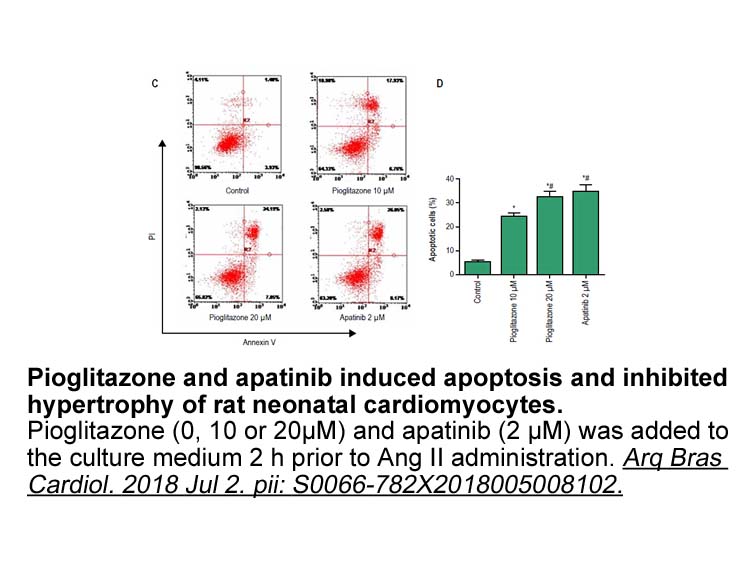
Conclusions In this paper we describe the recombinant expression and characterization of ParI, a C5-DNA-MTase from P. arcticus 273–4. To our knowledge, this is the first characterization of an orphan C5-DNA-MTase from a psychrophilic bacterium. The C5-DNA-MTase could not be expressed in regular E
-
Apart from neutrophil mediated inflammation the production o
2019-09-29
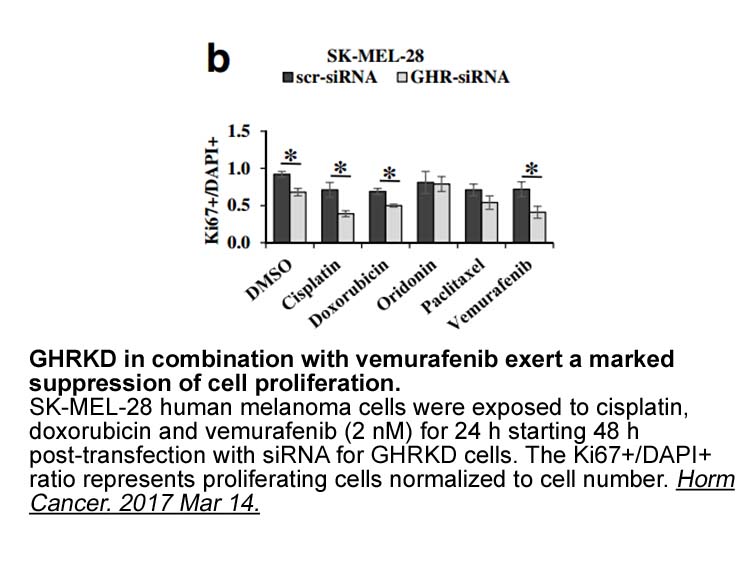
Apart from neutrophil-mediated inflammation, the production of pro-inflammatory cytokines and chemokines from PGD2/CRTH2 activated macrophages could further aggravate LPS-induced lung injury (Eguchi et al., 2011; Jandl et al., 2016). Interestingly, genetic ablation of CRTH2 diminishes the TNF-α prod
-
XMU-MP-1 As suggested by previous data we identified that
2019-09-29
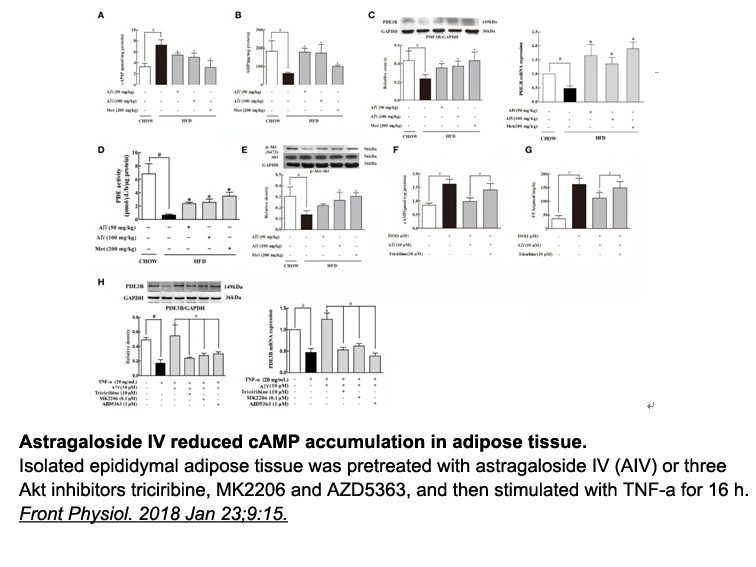
As suggested by previous data [45], we identified that the N-terminal and C-terminal domains of DDX3 were enriched in intrinsically disordered regions (Fig. 6). We also found that this characteristic was conserved in the external domains of several homologs of DDX3 (from yeast to human) and all desc
-
Unlike the CRF receptor the CRF a receptor binds
2019-09-29
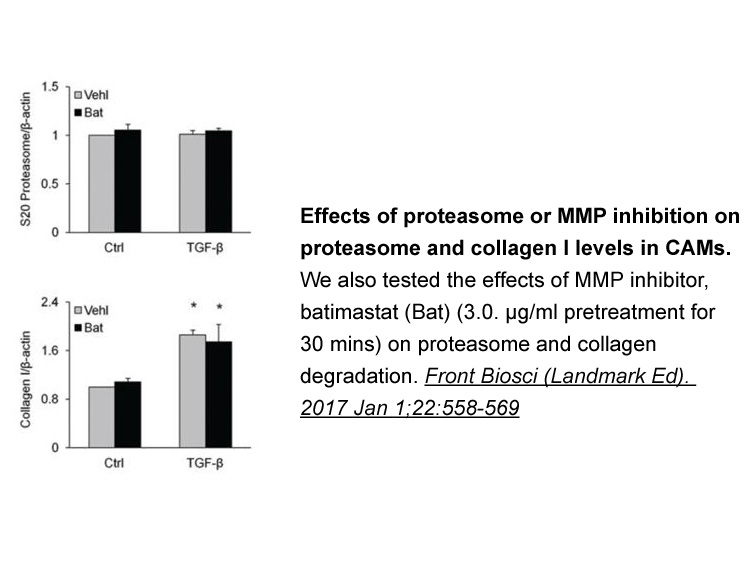
Unlike the CRF1 receptor, the CRF2(a) receptor binds and is activated by agonists with a broad range of potencies. Therefore, we assessed the ability of strong and weak ligands to desensitize retinoblastoma CRF2(a) receptors. Although stresscopin\'s N-terminus is two Cepharanthine longer than the N
-
gap-26 Distribution of DGTS in basidiomycete fungi has been
2019-09-29
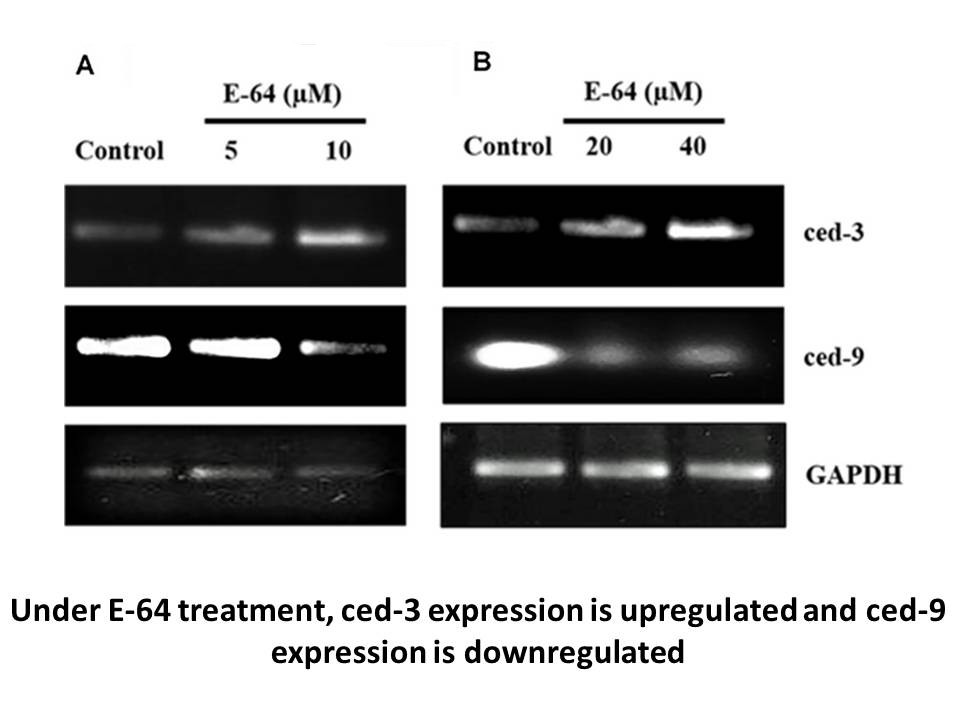
Distribution of DGTS in basidiomycete fungi has been demonstrated to be heterogeneous. In certain fungal taxons, such as Agaricales, Polyporales and Russulales, there are species that synthesize and species that do not synthesize DGTS that belong to the same order or even family (Dembitsky, 1996, Va
-
Population genetic summary statistics for five super
2019-09-29
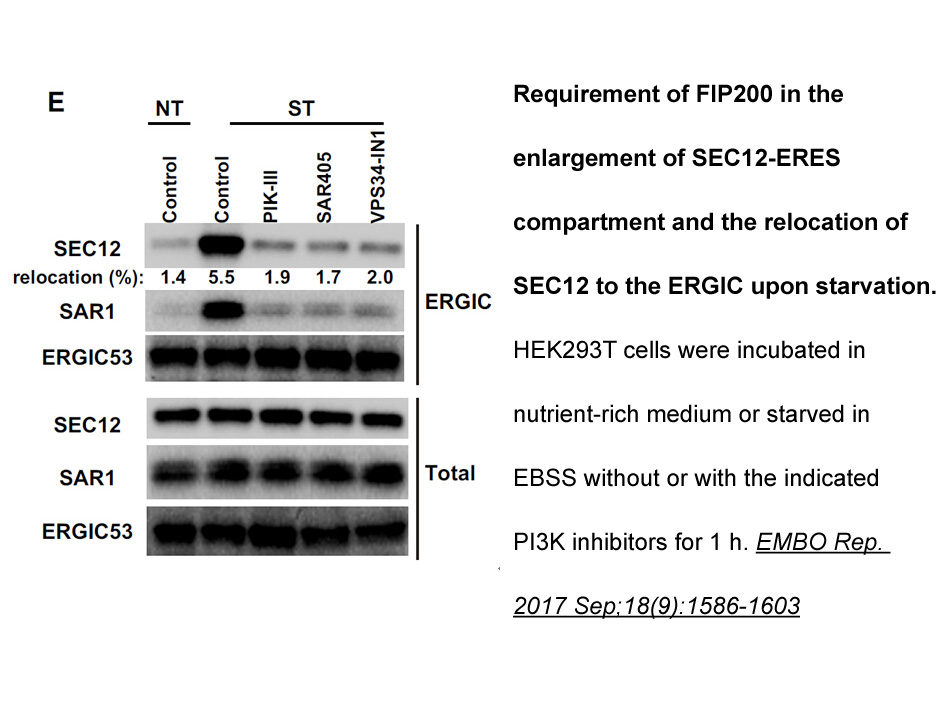
Population genetic summary statistics for five super- and 26 sub-populations, including haplotype and diplotype frequencies (analogous to allele and genotype frequencies), observed (Ho) and expected (He) heterozygosities, pairwise genetic distances, and tests for detection of departures from Hardy W
15827 records 1011/1056 page Previous Next First page 上5页 10111012101310141015 下5页 Last page